
")
Макарова Л.Е., Глянько А.К., Акимова Г.П., Соколова М.Г.
Бобово-ризобиальный симбиоз – уникальное биологическое явление, позволяющее бобовым растениям усваивать атмосферный азот с помощью эндогенного микросимбионта - клубеньковых бактерий, представителей семейства Rhizobiaceae. Известно, что начальные этапы взаимодействия растений с симбиотическими и патогенными микроорганизмами имеют много общего. Однако механизмы симбиотического взаимодействия являются более сложными и тонкими и недостаточно изученными. В связи с этим одним из важнейших аспектов проблемы развития бобово-ризобиального симбиоза является регуляция макросимбионтом начальных этапов симбиоза: проникновения ризобий в ткани корня и инициирования образования растительно-микробных структур – клубеньков.
Важную роль в регуляции процессов проникновения клубеньковых бактерий в ткани корня и образования клубеньков могут играть защитные реакции, сходные с начальными этапами взаимодействия растений с фитопатогенами. Бобово-ризобиальное взаимодействие, по-видимому, ведет у растения-хозяина к локально проявляющимся реакциям по репрессированию или индукции защитных механизмов, сопровождающихся генерацией активных форм кислорода и азота, усилением активности окислительных и других ферментов (пероксидаза, каталаза, полифенолоксидаза, супероксиддисмутаза), накоплением фенольных соединений, а также повышением активности антиоксидантной защиты, предотвращающей разрушительное действие свободных радикалов. Это требует глубоких переключений в метаболических системах растения, и, несомненно, должно происходить при участии фитогормонов, основных регуляторов жизнедеятельности растения. Появление локальных скоплений ИУК и ЦК, способствуют инициации митотических процессов в коре при формировании клубеньковых примордиев и они связаны с увеличением содержания фенольных соединений в клетках коры во время инфицирования ризобиями (Schultze, Kondorosi, 1998; Hirsch et al., 2001).
Изменения в фитогормональном балансе могут быть инициированы окислительными процессами. В частности, уровень и метаболизм ИУК взаимосвязаны с таким полифункциональным «стрессовым» ферментом как пероксидаза. Этот фермент не только окисляет ИУК, но и, как полагают, имеет центр специфического связывания с последней.
Все сказанное определило направления исследований сотрудников нашей лаборатории. Аспекты наших исследований отражены на схеме слайда. Они включают изучение изменений окислительного статуса, роли фитогормонов и роли эндогенных фенольных соединений в процессе развития реакции корневых клеток на инокуляцию ризобиями.
Комплекс проводимых нами исследований касается преимущественно начального периода формирования бобово-ризобиального симбиоза. В докладе представлена лишь небольшая часть полученных нами данных, подтверждающих существование защитных реакций в корнях и участие фитогормональной системы в указанный период взаимодействия симбионтов. Приводятся результаты, полученные в период, когда у корней проростков гороха обозначились различные по чувствительности к ризобиям зоны. Конкретно - через 1 сут после инфицирования этими бактериями.
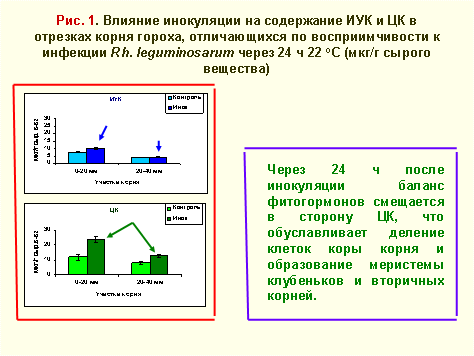
В указанный период взаимодействия симбионтов происходит целый ряд изменений в обмене корневых клеток растения-хозяина. Так, определение содержания ИУК и ЦК (рис.1) в двух различающихся по восприимчивости к ризобиальной инфекции участках от кончика корня растений гороха показало следующее. После инокуляции уровень ИУК увеличивался в основном в апикальном участке корня. Содержание этого фитогормона было низким и не изменялось после инокуляции во втором участке, примыкающем к первому. В обоих участках инокуляция вызывала повышение содержания ЦК и смещение баланса фитогормонов (ФГ) в сторону последних. Такое смещение баланса ФГ способствует меристематическим процессам в коре корня, связанным с образованием примордий клубеньков и вторичных корней.
Возрастанию уровня ИУК в апикальном участке корня, вероятно, способствует соответствующее снижение активности растворимой ПО и ИУКО (рис.2). Во втором участке достаточно высокие и лишь немного возрастающие после инокуляции уровни активности этих ферментов обусловливают низкое, практически не изменяющееся после инокуляции содержание ИУК.
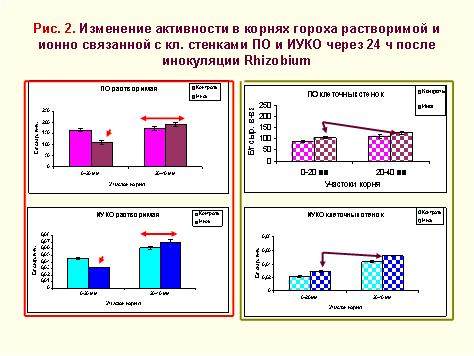
Небольшое увеличение активности ПО и ИУКО у инокулированных растений во фракции ионно-связанных с клеточными стенками, вероятно, вызвано процессами, происходящими в клеточной оболочке. В частности, ПО, как показано недавними работами, катализирует не только образование «перекрестных» связей в структурных белках, пектинах, лигнине, но и в инфекционных нитях, обеспечивающих дальнейший ход инфицирования растительных тканей.
Результаты определения активности растворимой и связанной форм ПО и ИУКО позволяют констатировать, что начальные этапы инфицирования корней гороха ризобиями сопровождаются перестройкой функционирования пероксидазной системы, обусловленной как компартментацией этих ферментов в клетках растений, так и изменением их функциональной активности.
Полученные данные дают основание полагать, что растворимая ПО участвует во взаимодействии растений с Rhizobium, по-видимому, через защитно-регуляторный механизм на уровне сигналинга, тогда как ПО клеточных стенок, очевидно, связана с механизмом регуляции, обеспечивающим, прежде всего, соответствующее состояние клеточной оболочки. Возрастание активности ПО в этом компартменте клетки можно считать показателем не только состоявшегося взаимодействия двух организмов, но и его определенного этапа, в частности - образования инфекционной нити.
При изучении физиологической роли активных форм кислорода (АФК) и азота (АФА) в защитно-регуляторных механизмах бобово-ризобиального симбиоза получены следующие результаты.
Прослеживание динамики изменения пероксида водорода и способности обезвреживания NO после инокуляции позволило установить, что ризобиальная инфекция способна регулировать интенсивность окислительного и нитрозативного стрессов в проростках гороха. Так, из рис. 3 видно, что инфицирование проростков ризобиями ведет через 30 мин. к резкому повышению содержания пероксида водорода в тканях корня, а в последующие экспозиции к его снижению до уровня контроля.
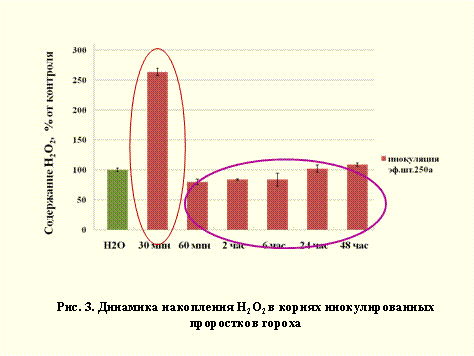
Влияние ризобиальной инфекции на содержание пероксида водорода в растении сходно с действием вирулентного фитопатогена, но не с действием авирулентного патогена, когда уровень пероксида водорода в тканях растения остается повышенным, независимо от длительности времени воздействия патогенна (Grant, Looke, 2000).
На рисунке 4 приведены данные, полученные методом микроскопии – по интенсивности флуоресценции вещества-зонда (4.5-диаминофлуоресцеин ацетат), реагирующего (после его деацилирования с помощью внутриклеточных эстераз) с NO с образованием флуоресцирующего зеленым светом соединения.
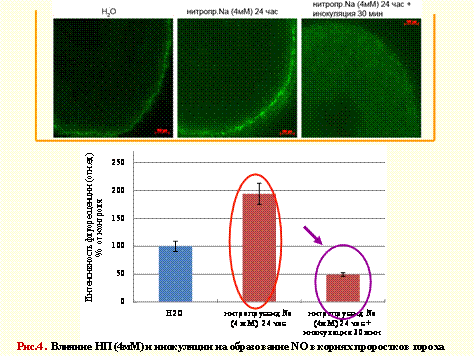
Из рис. следует, что ризобиальная инфекция способна снижать уровень оксида азота в одном из участков зоны адгезии корня проростков гороха (5-10 мм от кончика корня).
Из приведенных данных следует, что регуляция под влиянием ризобиальной инфекции уровня пероксида водорода и оксида азота, являющихся сигнальными и цитотоксическими компонентами клеток, свидетельствует об активных механизмах бобово-ризобиального симбиоза, могущих устранять «метаболические препятствия» на пути установления симбиоза.
Реакция корневых клеток на инфекцию различна не только в восприимчивых и невосприимчивых к ризобиальной инфекции зонах корня, но в разных участках восприимчивой зоны. Это подтверждают полученные нами данные по изучению особенности реакции клеток в 5-миллиметровых отрезках корней, последовательно нарезанных вдоль корня, начиная от его апекса.
На рисунке 5 показано примерное расположение зон корня, установленных с помощью светового микроскопа. Это зоны адгезии, зоны корневых волосков, подразделенные на участки в зависимости от этапа роста и развития корневых волосков и характера их взаимодействия с ризобиями. Устанавлены и неинфицируемые участки корня. Неинфицируемыми ризобиями являются области корня, прилегающие к основанию, где до инокуляции были зрелые корневые волоски и область, где они отсутствовали изначально, и, вероятно, апикальный, растущий участок корня.
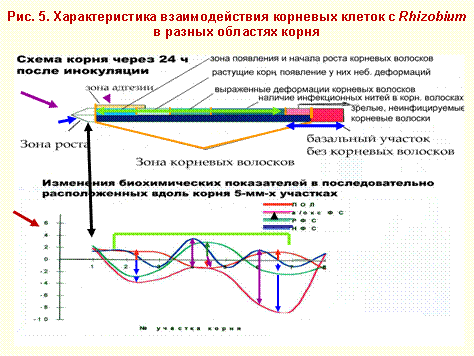
Приведенные ниже графики, характеризуют направленность и степень изменений в разных участках корней после их инокуляции биохимических показателей, характеризующих активность защитных систем клеток: ПОЛ, содержания «растворимых» липофильных фенольных соединений и их антиоксидантной активности в липидосодержащих системах, содержания фенольных соединений, связывающихся с макромолекулами клеточных стенок, то есть, участвующих в формировании физического барьера для ризобий.
Следует отметить альтернированный характер изменений перечисленных показателей вдоль корня.
Очевидно, клетки растущей части корня наиболее защищены от инфицирования ризобиями, поскольку все изученные нами показатели, имеющие отношение к защитным системам клеток, после инокуляции здесь возрастают. Добавим, что ранее в данной области корня сотрудниками нашей лаборатории отмечалось максимальное возрастание после инокуляции и уровня пероксида.
Характер изменения тех же показателей в разных участках зоны восприимчивости, включающей зону адгезии, деформаций корневых волосков и проникновения в них ризобий, неоднозначен. Так, в одних участках зоны восприимчивости, возрастание уровня ПОЛ сопряжено с заметным снижением содержания и существенным ослаблением антиоксидантной активности растворимых липофильных фенольных соединений. В некоторых участках этой зоны выявлено снижение уровня ПОЛ, которое, по-видимому, в основном обязано возрастанию концентрации в клетках тех же растворимых липофильных фенольных соединений со слабо выраженным снижением их антиоксидантной активности. Надо заметить, что само по себе повышение концентрации данных фенольных соединений можно рассматривать как проявление защитной реакции, поскольку возрастает опасность гибели для ризобий, случайно высвобожденных в цитоплазму растительных клеток из поврежденных каким-либо образом инфекционных нитей.
В некоторых областях зоны восприимчивости при ослаблении активности в ингибировании ПОЛ растворимых фенольных соединений усиливалось, хотя и в разной степени, связывание данных соединений в клеточных стенках, способствуя тем самым формированию физического барьера для ризобий.
Следовательно, механизмы реализации протекторных функций с участием ФС неодинаковы не только в разных по чувствительности к ризобиальной инфекции зонах корня, но и в разных участках чувствительной к данной инфекции зоны.
Итак, совокупность полученных нами данных свидетельствует об изменениях в метаболизме в ответ на инокуляцию. Они неодинаковы по активности и характеру проявлений в различающихся по восприимчивости к инфекции зонах корня.
Основываясь на приведенных данных можно констатировать, что ответ фитогормональной, прооксидантной и антиоксидантной защитных систем растения-хозяина на взаимодействие симбиотических партнеров представляет собой комплекс взаимосвязанных реакций, в результате которых сдерживается развитие опасных для клеток окислительных процессов, что дает возможность для нормального развития бобово-ризобиального симбиоза. Характер этого ответа в участках корня зависит от восприимчивости и характера взаимодействия его клеток с микросимбионтом.